| 30 of 30 |
| Brain Research | ||
| Volume 821, Issue 1 | SummaryPlus | |
| 6 March 1999 | Article | |
| Pages 241-249 | Journal Format-PDF (1782 K) |
PII: S0006-8993(99)01064-1
Copyright ©
1999 Elsevier Science B.V. All rights reserved
Interactive report
Hippocampal microtubule-associated protein-2 alterations with contextual
memory1
Nancy J. Woolfa,
*,
Marcus D. Zinnermanb
and Gail V. W. Johnsonb
a Laboratory of Chemical
Neuroanatomy, and Department of Psychology, UCLA, 405 Hilgard Ave., Los Angeles,
CA 90095-1563, USA
b Department of
Psychiatry and Behavioral Neurobiology, University of Alabama, Birmingham, AL
35294-0017, USA
Accepted 18 December 1998. Available online 3
March 1999. ![]()
Using immunohistochemistry and immunoblots, we show that alterations in hippocampal microtubule-associated protein-2 appear to be highly correlated with contextual memory as measured by significantly heightened fear responses. Compared to naive controls, rats trained in a novel context showed significantly increased immunostaining for the high molecular weight microtubule-associated protein-2a/b. This increase was observed 2 weeks after training and it was selective for hippocampal CA1 and CA2 pyramidal cells. Pre-exposure to the training context one month before training altered the hippocampal microtubule-associated protein-2 response; in these animals only the dentate gyrus showed significantly increased microtubule-associated protein-2a/b. Training-related increases in immunohistochemical staining for microtubule-associated protein-2 suggested that there was an increase in overall intact protein, an increase in immunoreactive breakdown products, or changes in protein compartmentalization. Immunoblots of hippocampal homogenates reacted with monoclonal antibodies to microtubule-associated protein-2a/b showed an increased presence of breakdown products in trained animals compared to untrained controls. Additional immunoblot studies demonstrated statistically significant decreases in the levels and/or phosphorylation state of the low molecular weight microtubule-associated protein-2c in the hippocampus of trained animals as compared to that of controls. These alterations in microtubule-associated protein-2 may reflect dendritic remodeling related to contextual memory storage.
Author Keywords: Dendritic remodeling; Fear conditioning;
Immunohistochemistry; Cytoskeletal protein; Immunoblot; Rat ![]()
Contextual, configural and spatial memory are among the few abilities in the rat that begin to approximate human memory. These kinds of memory are disrupted by lesions to the hippocampus [22, 28]. Analogous to the consolidation of human memory, the consolidation of contextual memory in the rat is not immediate; it takes between one and four weeks for contextual memory to become resilient to the amnesiac effects of hippocampal lesions [15].
Among the neural substrates related to memory, those involving microtubule-associated protein-2 (MAP-2) may signal dendritic remodeling [12]. MAP-2 is selectively localized to dendrites and has been proposed to play roles in neuritogenesis, synapse formation, and dendritic remodeling [12]. We have previously shown increased MAP-2 degradation and dendritic changes in auditory cortex following Pavlovian conditioning to a tone paired with an immediate shock [37]. Suggestive and definitive signs of MAP-2 degradation (i.e., increased immunohistochemistry and presence of breakdown products in immunoblots, respectively) were correlated with behavioral responding to the tone. These changes appeared in the auditory cortex immediately after 2¯3 days of training; however, there were no signs of MAP-2 degradation correlated with memory of the chamber in the hippocampus immediately after training.
To find if alterations in hippocampal MAP-2 occur during the consolidation of contextual memory we waited 2 weeks after 2 days of training and then assessed MAP-2 using both immunoblot and histochemical methods. MAP-2 consists of a pair of high molecular mass (280 kD) proteins, MAP-2a/b and a low molecular mass (70 kD) polypeptide, MAP-2c, which are generated by alternative splicing of a single gene [5, 17]. MAP-2 is extensively phosphorylated [31], and the phosphorylation state of MAP-2 modulates its function [4] and metabolism [11]. In this study, the monoclonal antibodies AP-14, AP-18, and HM-2 were used. AP-14 is a phosphate-independent antibody that recognizes MAP-2a/b, but not MAP-2c [2, 14]. AP-18 binds MAP-2a/b and c, but only when Ser136 is phosphorylated [1]. The specificity of these antibodies has been verified though reactions with purified protein, peptide mapping, and reactions with brain tissue [2, 3, 6] (see Ref. [25] for an outline of the methods used to test for specificity). Proteolytic fragments associated with MAP-2 are also recognized by these antibodies [2, 11, 13]. We also used the monoclonal antibody HM-2, which specifically reacts with the high molecular weight MAP-2 species [9].
The animals in this study were handled humanely and according to NIH guidelines. Sprague¯Dawley rats were bred on site in the UCLA Psychology Department vivarium to minimize stress due to transport. For the sake of uniformity, all rats were females, ranged 3¯4 months of age, and weighed 220¯250 g. Forty-eight rats were randomly assigned to training or control groups. Before training commenced, all rats were handled for 5 consecutive days. Animals were housed in individual cages before and after training. Naive rats were housed and handled in an identical manner, but did not receive training.
In experiment I, animals (n=24) were assigned to one of three groups. Two groups of animals were trained with a tone (2 KHz, 96 dB, 30 sec duration) and floor grid shock (1 mAmp, 1 sec duration) presented in a novel training chamber. After a 2 min adjustment period in the chamber, training commenced. One group of trained animals received the tone and then the shock immediately following. The second group received tone and shock unpaired (i.e., separated by 5 min to preclude tone conditioning in the presence of the conditioning stimuli as previously described [23, 37]). These two groups were trained for 2 consecutive days, and on each day they were given 4 trials (intertrial interval 10 min) as previously described [23, 37]. Each trial lasted approximately 40 min. The naive control group received no training or exposure to the chamber prior to testing.
Two weeks later, half the animals in the two trained groups and half the naive controls were tested by measuring freezing to the tone and to the training context. After a 2 min adjustment period, sampling of freezing behavior commenced. The observer, blind to the training, scored the presence or absence of freezing to the chamber during brief observations (4 observations per animal). Following a 2 min rest period, the observer similarly scored freezing behavior during the 30 sec tone (4 observations per animal). The animals were then removed from the training chamber (total testing time was approximately 5 min) and transcardially perfused within 1 h. Their brains were then processed for immunohistochemistry. (Comparisons between tested and untested animals showed that testing had no effect on subsequent immunohistochemical measures.)
In experiment II, animals (n=12) were assigned to one of the following groups: a group of animals pre-exposed to the chamber for 1 h one month before training and then trained with tone and shock paired, a group of animals trained with tone and shock paired (but not pre-exposed), or a group of naive animals (not trained or pre-exposed). The shock paired with tone training of these animals was the same as that described in experiment I. Brains from animals in experiment II were processed for immunohistochemistry.
In experiment III, animals (n=12) were assigned to one of the following groups: tone paired, tone unpaired or naive. These animals were trained in the same way as described for experiment I; however, the brains were processed biochemically. We did not do a study with pre-exposed animals and immunoblots.
Following the testing session (or on the day of testing for the untested animals), animals were deeply anesthetized with 70 mg/kg of pentobarbital and then sacrificed by transcardial perfusion with phosphate buffered saline (PBS) followed by 4% paraformaldehyde in 0.1 M phosphate buffer containing 0.2% picric acid (immunohistochemistry) or decapitated (biochemistry). Brains were then rapidly removed from the cranial cavity.
Protocols for immunohistochemistry and biochemistry were carried out as
previously described [11,
34,
37].
We used three antibodies to MAP-2: clones AP-14 and AP-18 (generous gift of Dr.
L.I. Binder) and HM-2 (Sigma Chemical Company, MO). The antibody 5H1 recognizing
![]() -tubulin and
-tubulin and ![]() -tubulin breakdown product was also
provided by L.I. Binder (for characterization see Ref. [33]).
-tubulin breakdown product was also
provided by L.I. Binder (for characterization see Ref. [33]).
Sectioning for immunohistochemistry was done at 40 ![]() m intervals through the entire
rostrocaudal extent of the hippocampus. Tissue sections were collected in PBS
and then incubated free-floating in vials containing primary antibodies diluted
in PBS (1:500). Tissue sections from differently trained animals were incubated
together in the same vial to avoid differences in the antibody concentration due
to the degree of binding to the tissue. After 24 h incubation in the primary
antibodies, tissue sections were incubated in biotinylated secondary antibodies
and avidin¯biotin (Vectastain; Burlingame, CA). Finally, the tissue
sections were reacted in a solution containing 0.05% diaminobenzidine, 0.01%
hydrogen peroxide, 2.5% nickel ammonium sulfate, and PBS. Control sections in
which secondary, but no primary, antisera were applied showed no staining.
m intervals through the entire
rostrocaudal extent of the hippocampus. Tissue sections were collected in PBS
and then incubated free-floating in vials containing primary antibodies diluted
in PBS (1:500). Tissue sections from differently trained animals were incubated
together in the same vial to avoid differences in the antibody concentration due
to the degree of binding to the tissue. After 24 h incubation in the primary
antibodies, tissue sections were incubated in biotinylated secondary antibodies
and avidin¯biotin (Vectastain; Burlingame, CA). Finally, the tissue
sections were reacted in a solution containing 0.05% diaminobenzidine, 0.01%
hydrogen peroxide, 2.5% nickel ammonium sulfate, and PBS. Control sections in
which secondary, but no primary, antisera were applied showed no staining.
The mounted tissue sections were analyzed by an Olympus microscope equipped with a Bioquant image analysis system. Microdensitometric measurements were made in the dorsal hippocampus (CA1, CA2, CA3, the dentate gyrus, and the subiculum) directly from tissue sections of each animal. These assessments were made blind to the training each animal received.
For the immunoblot study (experiment III), the entire hippocampus proper
(CA1, CA2, CA3, and DG) and parietal cortex from both sides of the brain was
rapidly dissected on ice. These samples were immediately frozen in dry ice.
Hippocampal and cortical samples were homogenized in 10 vol of sample buffer
(125 mM Tris-Cl, pH 6.8: 2% SDS, 10% glycerol and 5% ![]() -mercaptoethanol. After measuring
the protein concentration by acid precipitation, samples containing 80
-mercaptoethanol. After measuring
the protein concentration by acid precipitation, samples containing 80 ![]() g protein were diluted in the sample
buffer containing 0.02% bromophenol and run on 6¯12% gradient
SDS-polyacrylamide gels. Immunoblots were quantitated using BioRad Video
Densitometer 620.
g protein were diluted in the sample
buffer containing 0.02% bromophenol and run on 6¯12% gradient
SDS-polyacrylamide gels. Immunoblots were quantitated using BioRad Video
Densitometer 620.
We used ANOVA to statistically analyze all data. As a post-hoc test, we selected the Student-Newman¯Keuls test.
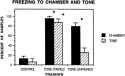
When placed in the training chamber 2 weeks after fear conditioning, trained animals exhibited very high levels of freezing; levels that were significantly increased above that of the extremely low levels exhibited by naive controls (see Fig. 1 for results from experiment I). Significantly increased freezing in the chamber was observed for trained animals regardless of whether the training involved tone followed immediately by shock (tone paired) or tone and shock spaced 5 min apart (tone unpaired). Although evidence of a contextual memory of the chamber appeared in both groups of animals trained in experiment I, only rats trained to the tone paired with shock demonstrated high levels of freezing to the tone, significantly above control levels (Fig. 1).
Fig. 1. Freezing in the chamber or to the tone as demonstrated 2 weeks after 2 days of training. Following a 2 min adjustment period, amounts of freezing were measured and are expressed here as the percent of samples. Four brief samples (freezing or not freezing) were taken for each animal at the onset of the 2 min exposure period to the training chamber and during the 30 sec exposure to the tone. Data from 4¯6 animals are averaged together for each measure. *Significantly increased above control values, Student-Newman¯Keuls post-hoc analyses, p<0.01.
In experiment II, pre-exposure to the chamber one month before animals were trained with tone and shock paired did not alter the high levels of freezing to the chamber or to the tone (measured 2 weeks after training). Freezing was recorded during 100% of the samples for animals in the pre-exposed group as compared to freezing occurring during 93.8% of samples for animals trained without pre-exposure. Comparatively, the naive animals in experiment II froze during 43.8% of the samples. The high levels of freezing exhibited by the trained animals in experiment II, regardless of pre-exposure, were similar to the high levels of freezing in animals trained with tone and shock paired in experiment I.
In experiment I, immunohistochemistry revealed differences between control animals and both groups of trained animals showing a memory for the context. As compared to tissue sections from controls (see Fig. 2A), tissue sections from both groups of trained animals that were incubated together in monoclonal antibody AP-14 displayed increased MAP-2a/b immunohistochemistry in CA1 (shown) and CA2 (not shown) of the hippocampus (see Fig. 2B and C). MAP-2a/b alterations were localized to apical dendrites and pyramidal somata, as exemplified in Fig. 3. Compared to the lighter staining pattern for MAP-2a/b in control animals (Fig. 3A and D), pyramidal cells and dendrites were more heavily stained in trained animals (Fig. 3B, C and E). An anticipated exception to this case was when animals were pre-exposed to the chamber; pre-exposure rendered pyramidal cell staining more like that of control animals (Fig. 3F). Training-related changes in MAP-2a/b were not observed outside the hippocampus. For example, no changes were observed in the parietal, occipital or temporal cortices surrounding the hippocampus. Similar results were obtained from sections stained with the antibody HM-2, which also recognizes MAP-2a/b.
Fig. 2. MAP-2 in the dorsal hippocampus at approximately 5.8 mm posterior to Bregma captured by a pseudocolor image on NIH 1.56 (red indicates greatest level of staining, yellow indicates low to medium levels, and blue the lowest levels). MAP-2a/b was recognized immunohistochemically by the AP-14 antibody. Tissue sections from differently trained animals were incubated together in the same vial and finally reacted with nickel-intensified diaminobenzidine. (A) Control animals showed mostly low to medium levels (yellow) of MAP-2a/b in CA1, whereas more intense (red) staining appeared in CA1 when animals retained a memory of the context due either to training with tone delivered paired with shock (B) or to training with tone delivered unpaired with shock (C). No reliable training-related differences appeared in the dentate gyrus (DG) or subiculum (S). Densitometric measurements and significance levels of training-related differences for CA1 and CA2 are found in the text. Scale bar equals 1 mm.
(149K)
Fig. 3. Hippocampal pyramidal cells immunostained for MAP-2 in CA1 of the hippocampus in animals sacrificed 2 weeks after 2 days of training. In naive control animals shown in (A) and (D), the large pyramidal cells and their apical dendrites (arrows) were lightly to moderately stained for MAP-2, in contrast to the moderate to intense staining for MAP-2 found in pyramidal cells and their apical dendrites in animals trained to tone paired with shock (B) and (E) or tone unpaired with shock (C). (Note that some cells and apical dendrites in tone paired with shock animals were also lightly stained: asterisk in frame (B).) Pre-exposure to the chamber one month before training resulted in lighter staining for MAP-2 (F), similar to that of control animals (D). These visual observations correspond with statistically significant differences in optical density measurements (see text). Frames (A), (B) and (C) illustrate immunohistochemical staining with HM-2, and frames (D), (E) and (F) illustrate immunostaining for MAP-2 with AP-14. Scale bar in B equals 20
m.
A completely different staining pattern was observed, however, in tissue sections reacted with AP-18, an antibody that specifically recognizes Ser136 phosphorylated MAP-2a/b, as well as MAP-2c. Overall, AP-18 staining of the hippocampus was markedly more intense than that of the surrounding neocortex and it was markedly more intense than hippocampal staining with AP-14 or HM-2. (Staining with AP-14 or HM-2 was not more intense in the hippocampus as compared to the neocortex.) Moreover, there were no obvious training-related differences in the hippocampus of sections reacted with AP-18.
Densitometric measurements of MAP-2a/b reacted with AP-14 or HM-2 confirmed the training-related changes observed in the hippocampus. Optical densities in CA1 and CA2 of animals receiving tone and shock paired (116.33±1.86) or tone unpaired with shock (102.67±5.78) were significantly (p<0.01) above that of controls (81.67±3.33). No significant training-related changes in MAP-2a/b were found for CA3 or the dentate gyrus in sections reacted with AP-14 or HM-2. In agreement with our general observations, no training-related differences were found in sections reacted with AP-18.
When hippocampal MAP-2a/b levels assessed with AP-14 or HM-2 were compared to the amount of freezing in the chamber for each subject, the two measures were significantly correlated (R=0.931; p<0.0005). This high correlation between MAP-2a/b and freezing in the chamber suggests a strong relationship between increased immunohistochemical staining for MAP-2a/b in the hippocampus and a fear-related memory of the chamber. (Fig. 4 shows data suggesting that this increase in MAP-2a/b immunohistochemistry may reflect elevated degradation of intact MAP-2a/b.)
Fig. 4. Immunoblots of hippocampal homogenates from control and trained rats probed with the phosphate-independent MAP-2a/b antibody: AP-14 (1:8000). No AP-14 reactive breakdown product (BDP) at ~90kD is present in the hippocampus of the control (C). However, the BDP is clearly evident in the hippocampal samples from rats trained in a novel chamber with tone paired with shock (TP) or tone unpaired with shock (TU). The positions at which molecular mass standards migrated are indicated at the left (kD).
In experiment II, pre-exposure of the animals to the training chamber one month before training inhibited the training-related response of MAP-2a/b in the hippocampus. Similar to the outcome of experiment I, those animals trained in experiment II that had not been pre-exposed to the training chamber showed significantly (p<0.05) increased AP-14 binding to MAP-2a/b in the dorsal hippocampal sectors CA1 and CA2 and no training-related effects in CA3, the dentate gyrus, or the subiculum. Following pre-exposure to the chamber one month earlier, training did not produce any significant alterations in CA1, CA2, CA3 or the subiculum; there was a significant (p<0.05) increase in MAP-2a/b in the dentate gyrus compared to naive controls, however.
In experiment III, immunoblot assays revealed training-related differences in
the appearance of breakdown products (for MAP-2a/b and ![]() -tubulin) and intact protein levels
(MAP-2c). The appearance of discrete MAP-2a/b breakdown products was distinctly
different in the trained versus control groups. Only one of the four controls,
for example, exhibited a 90 kD AP-14 immunoreactive band, and it was lightly
stained. This band at 90 kD was clearly present in two out of four samples from
the animals trained with tone unpaired with shock and three out of four animals
trained with tone paired with shock (Fig.
4). Results similar to those obtained with AP-14 were obtained in
immunoblots reacted with the monoclonal antibody HM-2. There was also a
significant increase (p<0.05) in an immunoreactive breakdown product
of
-tubulin) and intact protein levels
(MAP-2c). The appearance of discrete MAP-2a/b breakdown products was distinctly
different in the trained versus control groups. Only one of the four controls,
for example, exhibited a 90 kD AP-14 immunoreactive band, and it was lightly
stained. This band at 90 kD was clearly present in two out of four samples from
the animals trained with tone unpaired with shock and three out of four animals
trained with tone paired with shock (Fig.
4). Results similar to those obtained with AP-14 were obtained in
immunoblots reacted with the monoclonal antibody HM-2. There was also a
significant increase (p<0.05) in an immunoreactive breakdown product
of ![]() -tubulin in the tone paired
(0.455±0.020) and unpaired animals (0.443±0.070) as compared to the controls
(0.259±0.103). Levels of
-tubulin in the tone paired
(0.455±0.020) and unpaired animals (0.443±0.070) as compared to the controls
(0.259±0.103). Levels of ![]() -tubulin were not altered by the training (not shown).
-tubulin were not altered by the training (not shown).
MAP-2c was also altered in the hippocampus with the training that produced a contextual memory. As shown in Fig. 5, immunoblotting with AP-18 revealed that the reactive band representing MAP-2c was significantly (p<0.01) reduced in the tone paired and unpaired animals (0.173±0.009; 0.191±0.003) compared to naive controls (0.280±0.024). MAP-2c levels and the amount of freezing exhibited by individual animals were negatively correlated (R=-0.682; p<0.05); therefore, the decrease in MAP-2c was correlated with the fear response elicited by the chamber. No changes were found in the parietal cortex, which served as the control brain region in all the immunoblot studies.
Fig. 5. Immunoblots of hippocampal homogenates from control and trained rats probed with the phosphate-dependent MAP-2a/b antibody: AP-18. AP-18 recognizes MAP-2a/b and MAP-2c when Ser 136 is phosphorylated. Fifty micrograms of protein was run on each lane of a 7% SDS polyacrilamide gel and probed with AP-18 (1:4000). MAP-2c was significantly decreased in rats trained in a novel chamber with tone paired with shock (TP) or tone unpaired with shock (TU) compared to controls (C).
The behavioral response of an immobilized crouching posture was routinely elicited in previously trained animals when those animals were merely placed in the training chamber 2 weeks after training, whereas untrained animals rarely exhibited this response. Thus, a contextual memory was evident in all the trained animals. Across all animals, trained and untrained, conditioned fear of the context was highly correlated (p<0.0005) with MAP-2a/b immunohistochemical measures in CA1 and CA2 of the hippocampus. Conditioned fear of the tone was not observed in the tone unpaired with shock group, yet these animals showed MAP-2 changes in the hippocampus at 2 weeks after training that were similar to those of the tone paired with shock group. From these results, we can conclude that contextual memory, and not conditioning to tone, is affiliated with MAP-2 changes in CA1 and CA2 of the hippocampus.
Pre-exposure to the training context reduced the role that the dorsal hippocampus played in the processing of this contextual memory. Following pre-exposure to the context, MAP-2 immunohistochemistry in CA1 or CA2 of the hippocampus was not increased above control levels. Thus, this result is largely consistent with an earlier finding showing that pre-exposure to the context enables contextual fear conditioning following dorsal hippocampal lesions [38].
Training-related increases in MAP-2 immunohistochemistry found for CA1 and CA2 of the hippocampus were highly replicable. These increases above control levels were found for the two groups of trained animals (tone paired and tone unpaired) that exhibited high levels of behavioral responding to the chamber and with the two antibodies that selectively react with MAP-2a/b (AP-14 and HM-2). Moreover, increases in MAP-2 immunohistochemical staining were found for the control animals in experiment II that were trained but not pre-exposed to the training chamber. Immunoblot analyses using AP-14 revealed no significant differences in the levels of MAP-2a/b intact protein in the hippocampus of trained and untrained rats; thus, the increased immunohistochemical staining was not due to increased intact protein. Instead, an increased presence of breakdown products was evident in the immunoblots from trained animals compared to that of controls. Enhanced immunohistochemical staining occurs with the accumulation of highly immunoreactive breakdown products [29, 37]. Thus, elevated MAP-2 degradation appears to be the most likely interpretation of the increased immunohistochemical staining shown here.
Other data from the literature suggest that the degradation of high molecular
weight MAP-2a/b may be a component of long-term memory storage. For one,
long-term memory is interrupted by inhibitors of calpain [18]
and MAP-2a/b is particularly sensitive to calpain [13].
Incubation of purified MAP-2 with calpain in vitro produces a prominent AP-18
immunoreactive 90 kD breakdown product [13];
however, the 90 kD breakdown product of MAP-2a/b in the present study was only
detected with the phosphate-independent AP-14 and not by AP-18. Moreover, these
two MAP-2 breakdown products cannot be the same since AP-14 and AP-18 interact
with epitopes on opposite ends of the MAP-2 molecule. Nonetheless, it is
conceivable that calpain-mediated degradation of MAP-2 played a role in the
present study, but that the in vitro and in vivo cleavage sites are not
identical and hence different breakdown products are formed. It is not clear
what, if any, relationship exists between MAP-2 and the ![]() -tubulin breakdown products that
were increased in both groups of trained animals compared to the control group.
It is possible that tubulin degradation is part of the cytoskeletal remodeling
that occurs with memory consolidation.
-tubulin breakdown products that
were increased in both groups of trained animals compared to the control group.
It is possible that tubulin degradation is part of the cytoskeletal remodeling
that occurs with memory consolidation.
In the present study, MAP-2 alterations were commonly found within the apical dendrites of hippocampal pyramidal cells as a function of training. Van der Zee and colleagues [32] have similarly shown spatial learning is accompanied by altered apical dendrites of pyramidal cells in CA1 and CA2 using muscarinic receptor immunohistochemistry. The MAP-2 alterations reported here seem to represent breakdown of intact protein. The cytoskeletal matrix within segments of the apical dendrite or its branches may be degraded prior to new spines or branches being formed [35]. Degradation could be immediately followed by de novo synthesis of MAP-2, especially since MAP-2 can be synthesized locally within the dendrite [16]. Several previous findings suggest that changes in dendrite spines and branches may occur with contextual and spatial memory. Spatial learning in rats is accompanied by increased spine density on CA1 pyramidal cells [21]. Increased numbers of dendritic branches have not yet been detected in the hippocampus with spatial learning; however, maze learning has been shown to increase dendritic branching in the occipital cortex [7]. Hippocampal pyramidal cells do have the capacity to increase dendritic branch number; self-stimulation behavior produces this effect [26].
Tissue sections reacted with both AP-14 and HM-2 (selective for MAP-2a/b) invariably showed the same training-related increases in CA1 and CA2 above that of controls, whereas the histochemical specimens reacted with AP-18 (recognizing Ser136 phosphorylated MAP-2a/b and MAP-2c) showed no significant differences or trends between trained and control animals. The most parsimonious interpretation is that the different isoforms (MAP-2a/b versus MAP-2c) were differentially affected by training; hence only antibodies selective for MAP-2a/b would be expected to show the training-related increases and the result would be obscured in sections reacted with antibodies that additionally recognize MAP-2c. A further confounding factor is that the antibodies in question also differ in their selectivity for MAP-2 and MAP-2 fragments with respect to phosphorylation at the Ser136 site.
The immunoblot studies support the suggestion of isoform specific responses. In particular, the immunoblots revealed a decrease in MAP-2c with AP-18 in trained animals compared with controls. No decreases in intact MAP-2a/b were observed. The reason that the MAP-2c decrease was not observed histochemically is likely to be due to the fact that MAP-2c represents a relatively small portion of what is recognized by AP-18. No breakdown products were revealed in immunoblots with AP-18 further suggesting that the breakdown products accumulating in vivo are not phosphorylated at Ser136. Hence one would not expect to see the increased immunohistochemical response indicative of highly reactive breakdown products with AP-18.
Because the immunoblot studies of MAP-2c were carried out with the monoclonal antibody AP-18, which recognizes MAP-2 isoforms only when Ser136 is phosphorylated, the apparent decrease in MAP-2c could alternatively indicate a dephosphorylation of MAP-2c. There were no additional data indicating MAP-2c was degraded in the hippocampus of the trained rats; for example, there were no detectable immunoreactive products below intact MAP-2c in samples obtained from control or trained rats. However, potential breakdown products may not have been recognized by AP-18; also, further processing of the fragments may have occurred.
Without clear indications of MAP-2c breakdown, it is possible that MAP-2c was dephosphorylated with the training that produced contextual memory. Dephosphorylation of high molecular weight MAP-2 occurs, for example, in response to NMDA activation and results in potentiated responses in hippocampal slices, lasting up to tens of minutes [20, 24]. Dephosphorylation of MAP-2c could conceivably contribute to the contextual memory process through the mediation of an early temporal phase of potentiation. Long-term potentiation appears soon after birth in a sequence of phases that are added step-by-step during the early postnatal period [10, 30]. MAP-2c, which is especially abundant in the early postnatal period of development [19] is available to participate in the formation of the early temporal phases of potentiation.
While it is well established that MAP-2c is abundantly expressed in the developing brain [19] recent studies have detected substantial quantities of MAP-2c in the adult brain [8]. The role of MAP-2c in plasticity accompanying early development is well known [19] and may parallel its newly found role in memory formation in the adult hippocampus. Given the histochemical staining patterns obtained with AP-14 and AP-18, adult MAP-2c may be more abundant in the hippocampus than in the neocortex. Thus, adult MAP-2c may turn out to be particularly important to memory formation that relies on the hippocampus, such as remembering a training context. Still, developmental data suggest MAP-2a/b, rather than MAP-2c, is necessary for permanent structural changes essential to long-term configural memory. The developmental age when the high molecular weight isoform MAP-2a reaches adult levels, about 20 days of age [19], is exactly the same age that rat pups demonstrate the ability to form an enduring configural memory [27].
In conclusion, degradation and dephosphorylation of MAP-2 may represent
prerequisite steps leading to long-term memory storage [36].
The current results further suggest that the various isoforms of MAP-2 are
differentially affected during memory consolidation. Two weeks after
conditioning in a novel context, hippocampal MAP-2c is decreased or
dephosphorylated at its N-terminus. MAP-2a/b appears to be degraded in the
hippocampus of trained animals as compared to controls, and those MAP-2a/b
degradation products do not appear to contain a phosphorylated Ser136 site. ![]() References
References
1. B. Berling, H. Wille, B. Roll, E.M. Mandelkow, C. Garner and E. Mandelkow, Phosphorylation of microtubule-associated proteins MAP2a,b and MAP2c at Ser136 by proline-directed kinases in vivo and in vitro. Eur. J. Cell Biol. 64 (1994), pp. 120¯130. Abstract
2. L.I. Binder, A. Frankfurter and L.I. Rebhun, Differential localization of MAP-2 and tau in mammalian neurons in situ. Ann. N.Y. Acad. Sci. 466 (1986), pp. 145¯166. EMBASE
3.
L.I. Binder, M.R. Payne, H. Kim, V.R. Sheridan, D.K. Schroeder, C.C. Walker and
L.I. Rebhun, Production and analysis of monoclonal hybridoma antibodies specific
for ![]() -tubulin and MAP2. J.
Cell Biol. 95 (1982), p. 349a.
-tubulin and MAP2. J.
Cell Biol. 95 (1982), p. 349a.
4. B. Brugg and A. Matus, Phosphorylation determines the binding of microtubule-associated protein 2 (MAP2) to microtubules in living cells. J. Cell Biol. 114 (1991), pp. 735¯743. EMBASE
5. W.J. Chung, S. Kindler, C. Seidenbecher and C.C. Garner, MAP2a, an alternatively spliced variant of microtubule-associated protein 2. J. Neurochem. 66 (1996), pp. 1273¯1281.
6. C.G. Dotti, G.A. Banker and L.I. Binder, The expression and distribution of the microtubule-associated proteins tau and microtubule-associated protein 2 in hippocampal neurons in the rat in situ and in cell culture. Neuroscience 23 (1987), pp. 121¯130.
7. W.T. Greenough, J.M. Juraska and F.R. Volkmar, Maze training effects on dendritic branching in occipital cortex of adult rats. Behav. Neural. Biol. 26 (1979), pp. 287¯297. EMBASE
8. S. Halpain and G. Withers, A phosphorylation state-dependent antibody defines isoforms of MAP2 with distinctive subcellular localization and developmental expression. Soc. Neurosci. Abstr. 21 (1995), p. 800.
9. G. Huber and A. Matus, Differences in the cellular distributions of two microtubule-associated proteins, MAP1 and MAP2, in rat brain. J. Neurosci. 4 (1984), pp. 51¯60.
10. P.S. Jackson, T. Suppes and K.M. Harris, Stereotypical changes in the pattern and duration of long-term potentiation expressed at postnatal days 11 and 15 in the rat hippocampus. J. Neurophysiol. 70 (1993), pp. 1412¯1419. Abstract
11. G.V.W. Johnson and V.G. Foley, Calpain-mediated proteolysis of microtubule-associated protein 2 (MAP-2) is inhibited by phosphorylation by cAMP-dependent protein kinase, but not by Ca2+/calmodulin-dependent protein kinase II. J. Neurosci. Res. 34 (1993), pp. 642¯647. EMBASE
12. G.V.W. Johnson and R.S. Jope, The role of microtubule-associated protein 2 (MAP-2) in neuronal growth, plasticity, and degeneration. J. Neurosci. Res. 33 (1992), pp. 505¯512. EMBASE
13. G.V.W. Johnson, J.M. Litersky and R.S. Jope, Degradation of microtubule-associated protein 2 and brain spectrin by calpain: a comparative study. J. Neurochem. 56 (1991), pp. 1630¯1638. EMBASE
14. N. Kalcheva, J.S. Albala, L.I. Binder and B. Shafit-Zagardo, Localization of specific epitopes on human microtubule-associated protein 2. J. Neurochem. 63 (1994), pp. 2336¯2341. Abstract
15. J.J. Kim and M.S. Fanselow, Modality-specific retrograde amnesia of fear. Science 256 (1992), pp. 675¯677. EMBASE
16. S. Kindler, R. Muller, W.J. Chung and C.C. Garner, Molecular characterization of dendritically localized transcripts encoding MAP2. Mol. Brain Res. 36 (1996), pp. 63¯69. Abstract | Journal Format-PDF (600 K)
17. S. Kindler, B. Schulz, M. Goedert and C.C. Garner, Molecular structure microtubule-associated protein 2b and 2c from rat brain. J. Biol. Chem. 265 (1990), pp. 19679¯19684. BIOTECHNOBASE EMBASE
18. G. Lynch and M. Baudry, The biochemistry of memory: a new and specific hypothesis. Science 224 (1984), pp. 1057¯1063. EMBASE
19. A. Matus, Microtubule-associated proteins: their potential role in determining neuronal morphology. Annu. Rev. Neurosci. 11 (1988), pp. 29¯44.
20. R.J. Montoro, J. Diaz-Nido, J. Avila and J. Lopez-Barneo, N-methyl-D-aspartate stimulates the dephosphorylation of the microtubule-associated protein 2 and potentiates excitatory synaptic pathways in the rat hippocampus. Neuroscience 54 (1993), pp. 859¯871. EMBASE
21. M.B. Moser, M. Trommald and P. Andersen, An increase in dendritic spine density on hippocampal CA1 pyramidal cells following spatial learning in adult rats suggests the formation of new synapses. Proc. Natl. Acad. Sci. USA 91 (1994), pp. 12673¯12675. EMBASE BIOTECHNOBASE Elsevier BIOBASE
22. L. Nadel, The hippocampus and space revisited. Hippocampus 1 (1992), pp. 221¯229.
23. J.D. Oh, R.H. Edwards and N.J. Woolf, Choline acetyltransferase mRNA plasticity with Pavlovian conditioning. Expl. Neurol. 140 (1996), pp. 95¯99. Abstract | CrossRef
24. E.M. Quinlan and S. Halpain, Postsynaptic mechanisms for bidirectional control of MAP2 phosphorylation by glutamate receptors. Neuron 16 (1996), pp. 357¯368. EMBASE Elsevier BIOBASE
25. M.R. Payne, Monoclonal antibodies to contractile proteins, in: R.M. Dowben, J.W. Shay (Eds.), Cell and Muscle Motility, Plenum Press, New York, 1982, pp. 137¯190.
26. B.S. Rao, T. Desiraju and T.R. Raju, Neuronal plasticity induced by self-stimulation rewarding experience in rats ¯¯ a study on alteration in dendritic branching in pyramidal neurons of hippocampus and motor cortex. Brain Res. 627 (1993), pp. 216¯224.
27. J.W. Rudy, Contextual conditioning and auditory cue conditioning dissociate during development. Behav. Neurosci. 107 (1993), pp. 887¯891. Abstract
28. J.W. Rudy and R.J. Sutherland, The hippocampal formation is necessary for rats to learn and remember configural discriminations. Behav. Brain Res. 34 (1989), pp. 97¯109. EMBASE
29. P. Seubert, Y. Nakagawa, G. Ivy, P. Vanderklish, M. Baudry and G. Lynch, Intrahippocampal colchicine injection results in spectrin proteolysis. Neuroscience 31 (1989), pp. 195¯202. EMBASE
30. T.J. Teyler, A.T. Perkins and K.M. Harris, The development of long-term potentiation in hippocampus and neocortex. Neuropsychologia 27 (1989), pp. 31¯39. EMBASE
31. S. Tsuyama, Y. Terayama and S. Matsuyama, Numerous phosphates of microtubule-associated protein 2 in living rat brain. J. Biol. Chem. 262 (1987), pp. 10886¯10892.
32. E.A. van der Zee, J.C. Compaan, B. Bohus and P.G. Luiten, Alterations in the immunoreactivity for muscarinic acetylcholine receptors and colocalized PKC gamma in mouse hippocampus induced by spatial discrimination learning. Hippocampus 5 (1995), pp. 349¯362.
33. Y. Wang, P.A. Loomis, R.P. Zinkowski and L.I. Binder, A novel tau transcript in cultured human neuroblastoma cells expressing nuclear tau. J. Cell Biol. 121 (1993), pp. 57¯67.
34. N.J. Woolf, Cholinoceptive cells in rat cerebral cortex: somatodendritic immunoreactivity for muscarinic receptor and cytoskeletal proteins. J. Chem. Neuroanat. 6 (1993), pp. 375¯390. Abstract
35. N.J. Woolf, Global and serial neurons form a hierarchically arranged interface proposed to underlie memory and cognition. Neuroscience 74 (1996), pp. 625¯651. SummaryPlus | Article | Journal Format-PDF (945 K)
36. N.J. Woolf, The critical role of cholinergic basal forebrain neurons in morphological change and memory encoding: a hypothesis. Neurobiol. Learn. and Memory 66 (1996), pp. 258¯266. EMBASE Elsevier BIOBASE | CrossRef
37. N.J. Woolf, S.L. Young, G.V.W. Johnson and M.S. Fanselow, Pavlovian conditioning alters cortical microtubule-associated protein-2. Neuroreport 5 (1994), pp. 1045¯1048. Abstract
38.
S.L. Young, D.L. Bohenek and M.S. Fanselow, NMDA processes mediate anterograde
amnesia of contextual fear conditioning induced by hippocampal damage:
Immunization against amnesia by context preexposure. Behav. Neurosci.
108 (1994), pp. 19¯29. Abstract
![]()
1Published on the World Wide Web on 28 January 1999.
*Corresponding
author. Tel. : +1-310-206-7874; Fax: +1-818-760-2694; E-mail: nwoolf@ucla.edu ![]()
| Brain Research | SummaryPlus
Article Journal Format-PDF (1782 K) | |
| Volume 821, Issue 1 | ||
| 6 March 1999 | ||
| Pages 241-249 |
| 30 of 30 |
Send feedback
to ScienceDirect
Software and compilation © 2001 ScienceDirect. All rights
reserved.
ScienceDirect® is an Elsevier Science B.V. registered
trademark.
.gif)
.gif)