|
|
The Journal of Neuroscience, February 1, 2001,
21(3):1047-1055
1 Department of Psychology and 2 Brain Research Institute, University of California at Los Angeles, Los Angeles, California 90095, and 3 Department of Pharmacology, Texas Tech University Health Sciences Center, Lubbock, Texas 79430
| |
ABSTRACT |
|---|
We report here a series of experiments establishing a role for nerve growth factor and its high-affinity receptor TrkA in contextual memory consolidation. In all experiments, we trained rats in a novel chamber using tone and shock. Our first experiment revealed that endogenous nerve growth factor (NGF) increases in the hippocampus at a critical time during consolidation that occurs 1 week after training. NGF levels at other intervals (24 hr and 2 and 4 weeks after training) did not differ from those of naive control animals. In our second experiment, we blocked effects that NGF has at 1 week after training by infusing antisense TrkA phosphorothioate DNA oligonucleotide. Reduction of septohippocampal TrkA receptor expression selectively impaired memory consolidation for context but not for tone. Animals with antisense TrkA oligonucleotide infused into the medial septal area or CA1 of the hippocampus froze less when placed in the training chamber than did animals infused with inactive randomized oligonucleotide. At 4 weeks after training, antisense TrkA oligonucleotide had no effect on freezing. Third, we correlated levels of freezing with choline acetyltransferase (ChAT) and vesicular acetylcholine transporter (VAChT) immunohistochemistry. Antisense TrkA infused into CA1 of the hippocampus reduced cell body cross-sectional area for cholinergic cells in the medial septal area and decreased the density of hippocampal terminals labeled for ChAT and VAChT proteins. Cholinergic cell body measurements were significantly correlated with freezing. Taken together, these results indicate a role for nerve growth factor acting via the TrkA receptor on ChAT and VAChT proteins in contextual memory consolidation.
Key words: neurotrophins; Pavlovian conditioning; neural plasticity; hippocampus; DNA oligonucleotides; choline acetyltransferase; vesicular acetylcholine transporter
| |
INTRODUCTION |
|---|
Neurotrophins participate in plastic processes in the brain, including those
that accompany memory consolidation. Nerve growth factor (NGF) plays
a role in memory and attention tasks that rely on the cholinergic
septohippocampal pathway. NGF deprivation impairs spatial learning,
for example, and causes cholinergic hypofunction in the medial septal
area (Van Der Zee et al., 1995![]() ). Infusions
of NGF facilitate spatial memory performance and restore cholinergic
activity in aged and experimentally impaired animals (Janis et
al., 1995
). Infusions
of NGF facilitate spatial memory performance and restore cholinergic
activity in aged and experimentally impaired animals (Janis et
al., 1995![]() ; Fischer et al.,
1996
; Fischer et al.,
1996![]() ; Markowska et
al., 1996
; Markowska et
al., 1996![]() ).
Endogenous NGF levels also correlate with an animal's capacity for
spatial learning (Henriksson et al., 1992
).
Endogenous NGF levels also correlate with an animal's capacity for
spatial learning (Henriksson et al., 1992![]() ; Katoh-Semba and Kato,
1994
; Katoh-Semba and Kato,
1994![]() ).
).
A recent study further showed a positive relationship between NGF-induced
enhancement of memory and increases in cholinergic markers and
cholinergic cell body area (Gustilo et al., 1999![]() ).
Hippocampal NGF and NGF mRNA levels also increase after animals
are exposed to an enriched environment (Torasdotter et al., 1998
).
Hippocampal NGF and NGF mRNA levels also increase after animals
are exposed to an enriched environment (Torasdotter et al., 1998![]() ; Pham et
al., 1999b
; Pham et
al., 1999b![]() ). Because
enriched environments increase learning in a general way, one might
expect to find specific increases in NGF after specific learning
experiences.
). Because
enriched environments increase learning in a general way, one might
expect to find specific increases in NGF after specific learning
experiences.
We predicted that fluctuations in NGF accompanying memory consolidation of a
spatial context might occur in the hippocampus slightly before
structural changes. We found previously that signs of structural
plasticity (i.e., cytoskeletal protein degradation) appear in the
hippocampus 2 weeks after animals receive training that produces
contextual memory (Woolf et al., 1999![]() ). In the present
study, we used the same contextual memory task that was used in
our previous study and evaluated NGF levels at time intervals
ranging from 24 hr to 4 weeks after training.
). In the present
study, we used the same contextual memory task that was used in
our previous study and evaluated NGF levels at time intervals
ranging from 24 hr to 4 weeks after training.
Next, we tested whether NGF and expression of its high-affinity receptor TrkA
are essential for consolidating memory of a spatial context. We
infused antisense TrkA phosphorothioate DNA oligonucleotides into the
medial septal area and into CA1 of the hippocampus. It has been
established that antisense TrkA oligonucleotide reduces the synthesis
of TrkA receptor protein (Taglialatela et al., 1996![]() ).
).
Choline acetyltransferase (ChAT) and vesicular acetylcholine transporter
(VAChT) represent two known cholinergic markers (Woolf, 1991![]() ; Butcher et al.,
1992
; Butcher et al.,
1992![]() ; Bejanin et
al., 1994
; Bejanin et
al., 1994![]() ; Erickson
et al., 1994
; Erickson
et al., 1994![]() ; Roghani et al.,
1994
; Roghani et al.,
1994![]() ). In
cholinergic presynaptic terminals, ChAT synthesizes acetylcholine
(ACh), and VAChT is responsible for transport of ACh into synaptic
vesicles for regulated exocytotic release (Parsons et al., 1993
). In
cholinergic presynaptic terminals, ChAT synthesizes acetylcholine
(ACh), and VAChT is responsible for transport of ACh into synaptic
vesicles for regulated exocytotic release (Parsons et al., 1993![]() ). The VAChT gene is
localized to the first intron of the ChAT gene, and the expression of
these two genes is coregulated (Bejanin et al., 1994
). The VAChT gene is
localized to the first intron of the ChAT gene, and the expression of
these two genes is coregulated (Bejanin et al., 1994![]() ; Erickson et al.,
1994
; Erickson et al.,
1994![]() ; Berrard et
al., 1995
; Berrard et
al., 1995![]() ; Berse and
Blusztajn, 1995
; Berse and
Blusztajn, 1995![]() ; Usdin et
al., 1995
; Usdin et
al., 1995![]() ). Via the TrkA
receptor, NGF is known to upregulate ChAT and VAChT (Tian et al.,
1996
). Via the TrkA
receptor, NGF is known to upregulate ChAT and VAChT (Tian et al.,
1996![]() ; Takei et
al., 1997
; Takei et
al., 1997![]() ).
).
To establish effects on cholinergic cells and their markers, we assessed ChAT and VAChT expressions after TrkA antisense infusion. We also evaluated correlations between behavioral responses and ChAT- and VAChT-immunoreactive cell body cross-sectional area in the medial septal area and assessed terminal fields in the hippocampus.
Parts of this paper have been published previously in abstract form (Milov et
al., 1997![]() ; Woolf et
al., 1998
; Woolf et
al., 1998![]() ).
).
| |
MATERIALS AND METHODS |
|---|
Antisense oligonucleotides. We used two commercially available
synthetic DNA oligonucleotides: antisense TrkA oligonucleotide and a
randomized control oligonucleotide. Both oligonucleotides were fully
phosphorothioated. Antisense oligonucleotides directed to TrkA and
randomized controls have been designed and manufactured by Biognostik
(Göttingen, Germany). The antisense TrkA sequence
(5'-AACTGTTGTTGTGTCC-3') corresponds to nucleotides 623-638 of
rat TrkA (Meakin et al., 1992![]() ). A random
oligonucleotide (5'-GTAAATTGACCAGGAG-3') was used as a control. We
dissolved these oligonucleotides in a buffer containing10 mM Tris-Cl, pH 7.2, and 1 mM EDTA
to a final concentration of 200 µM.
). A random
oligonucleotide (5'-GTAAATTGACCAGGAG-3') was used as a control. We
dissolved these oligonucleotides in a buffer containing10 mM Tris-Cl, pH 7.2, and 1 mM EDTA
to a final concentration of 200 µM.
Animals. Seventy-two female Sprague Dawley rats were subjects in the present experiments. These animals were humanely treated according to both National Institutes of Health and University of California at Los Angeles (UCLA) guidelines. Three-month-old rats bred at UCLA (weight, 220-250 gm) were selected, at which time they were removed from group cages, placed in individual stainless steel cages, handled daily, and fed ad libitum. A 12/12 hr light/dark cycle was maintained, and the animals were trained and tested during the light cycle.
Behavioral training and testing. For experiment 1, we randomly assigned rats (n = 20) to one of five conditions: 0 (24 hr), 1, 2, and 4 week training-testing interval groups or a naive control group (see Table 1). Training consisted of placing the animals in a novel conditioning chamber where they received four tone-shock pairings (a 30 sec, 96 dB, 2 kHz tone and a 1 sec, 1 mA grid shock) with an intertrial interval of 10 min. Training continued for 2 consecutive days, and the different groups of trained rats were then tested at 24 hr to 4 weeks after 2 d of training (Table 1). One group of naive control rats was also tested. An observer blind to each animal's condition scored behavior from videotapes. Periods of immobility or freezing were measured during a 30 sec period starting 2 min after the rats were placed in the chamber. Two minutes after the onset of the first observation period, the tone came on, and freezing was measured during the 30 sec tone duration. Rats were killed 4 hr after testing, when the effects of testing had worn off.
|
To test for interactions between testing and training, we assigned rats (n = 16) to one of four conditions: trained and tested, trained and untested, untrained and tested, or untrained and untested. Training and testing were the same as above, except that the interval between training and testing was held constant at 1 week. We killed these rats within 1 hr after testing, when the effects of testing were in place.
In experiment 2, rats (n = 36) were randomly assigned to receive randomized control oligonucleotide or antisense TrkA oligonucleotide at two intervals after training (see Table 2). First, we trained these animals as described above for experiment 1. Next, the animals received surgical infusions of oligonucleotide either 1 or 4 weeks after training. Oligonucleotides were infused into the medial septal area (1 and 4 week groups) or CA1 of the hippocampus (1 week group only). We tested and killed animals 24 hr after infusion, except for one hippocampal group that was infused, tested, and killed 48 hr later.
|
ELISA measurements for NGF. Deeply anesthetized rats (injected with
90 mg/kg Nembutal, i.p.) were decapitated by guillotine. We then
rapidly dissected and saved the hippocampus from both sides of each
brain. We also saved the parietotemporal cortex as a reference brain
area. Next, brain samples were homogenized in an extraction buffer
containing 100 mM Tris-Cl, pH
7.0, 400 mM NaCl, 20 mM CaCl2, 1 mM PMSF,
4 mM EDTA, and 7 µg/ml aprotinin, and
then the mixtures were spun at 40,000 rpm for 10 min at 4°C.
We used the sandwich enzyme immunoassay technique to determine
NGF levels (anti-NGF and standards were purchased from Roche
Molecular Biochemicals, Indianapolis, IN). The supernatants were
collected, and 100 µl of each sample was pipetted into a well of
a 96-well microtiter plate (Immulon 2; Dynex, West Sussex, United
Kingdom) precoated with primary anti-NGF. After an overnight
incubation at 4°C, these plates were rinsed, then secondary
anti-NGF-![]() -galactosidase was applied, and the mixture was left
overnight at 4°C. Subsequently, the substrate chlorophenol-
-galactosidase was applied, and the mixture was left
overnight at 4°C. Subsequently, the substrate chlorophenol-![]() -D-galactopyranoside in a buffer
containing 100 mM HEPES, pH
7.0, 150 mM NaCl, 2 mM
MgCl2, and 1% bovine serum albumin was applied for
20 min. The microtiter plates were read on a V-Max plate reader
(Molecular Devices, Sunnyvale, CA) equipped with a 575 nm
filter.
-D-galactopyranoside in a buffer
containing 100 mM HEPES, pH
7.0, 150 mM NaCl, 2 mM
MgCl2, and 1% bovine serum albumin was applied for
20 min. The microtiter plates were read on a V-Max plate reader
(Molecular Devices, Sunnyvale, CA) equipped with a 575 nm
filter.
Infusion of antisense oligonucleotides. To anesthetize the rats, we
injected 65-70 mg/kg Ketamine, along with 8.6 mg/kg xylazine,
administered intraperitoneally. We then prepared rats for surgery
and placed them in the stereotaxic instrument. The medial septal
area (n = 20) or CA1 of the hippocampus (see below) was
targeted. Stereotaxic coordinates for the medial septal area (the
medial septal nucleus and vertical diagonal band nucleus) were set
at 0.5 mm anterior and 0.5 mm lateral to bregma and
6.0 and 7.5 mm ventral to the surface of the skull
according to the atlas of Paxinos and Watson (1997)![]() . At each brain site,
we infused 5 µl of oligonucleotide with a Hamilton microliter
syringe. The first injections we made were unilateral (two injection
sites at two depths, totaling 10 µl), after which we made
bilateral injections (four injection sites, totaling 20 µl).
. At each brain site,
we infused 5 µl of oligonucleotide with a Hamilton microliter
syringe. The first injections we made were unilateral (two injection
sites at two depths, totaling 10 µl), after which we made
bilateral injections (four injection sites, totaling 20 µl).
Because TrkA expression is maximally suppressed by antisense after 24 hr
(Taglialatela et al., 1996![]() ), we waited 24 hr
and then tested and immediately killed the animals. We tested
for tone and context-related memory by measuring freezing as
described for experiment 1. After the testing session, animals
were deeply anesthetized (90 mg/kg Nembutal, i.p.) and
transcardially perfused.
), we waited 24 hr
and then tested and immediately killed the animals. We tested
for tone and context-related memory by measuring freezing as
described for experiment 1. After the testing session, animals
were deeply anesthetized (90 mg/kg Nembutal, i.p.) and
transcardially perfused.
Additional animals (n = 16) had antisense TrkA
oligonucleotide and randomized oligonucleotide infused into CA1 of the
hippocampus. The stereotaxic coordinates used to approach the dorsal
CA1 of the hippocampus were 4.5 mm posterior and 3.0 mm
lateral to bregma and 3.0 mm ventral to the surface of the
skull. We infused a 5 µl oligonucleotide sample into each side
of the hippocampus. The first half of these animals were tested
24 hr after surgery, and the second half were tested 48 hr
later. We waited 48 hr in the latter case because we were
interested in retrograde effects on the cell bodies of medial septal
neurons. On the basis of studies using fluorescent retrograde
tracers, it takes 48 hr for medial septal neurons to
retrogradely transport maximal quantities of materials deposited in
the hippocampus (Woolf et al., 1984![]() ).
).
Immunohistochemistry and tissue analyses. We processed brain tissue
from animals in experiment 2 for immunohistochemistry according
to previously described protocols (Oh et al., 1991![]() ; Roghani et
al., 1998
; Roghani et
al., 1998![]() ). Briefly,
animals were deeply anesthetized and transcardially perfused with
50 ml of PBS followed by 250 ml of 4% paraformaldehyde in
0.1 M phosphate buffer containing 0.2%
picric acid at 4°C. We cut the brains at 40 µm intervals through
the rostrocaudal extent of the medial septal area in the frontal
plane of Paxinos and Watson (1997)
). Briefly,
animals were deeply anesthetized and transcardially perfused with
50 ml of PBS followed by 250 ml of 4% paraformaldehyde in
0.1 M phosphate buffer containing 0.2%
picric acid at 4°C. We cut the brains at 40 µm intervals through
the rostrocaudal extent of the medial septal area in the frontal
plane of Paxinos and Watson (1997)![]() . All tissue
sections through the medial septal nucleus and vertical diagonal band
nucleus were saved, starting with 1.6 mm anterior to bregma
through 0.3 mm posterior to bregma (anatomical landmarks were
used to determine anterior-posterior level). One-third of these
tissue sections were immunostained for ChAT, one-third were
immunostained for VAChT, and one-third were immunostained for TrkA.
We also saved sections through the dorsal CA1 region of the
hippocampus from animals receiving infusions in that site. Tissue
sections from differently trained animals were nicked at a
predetermined place on the cortex (for later identification) and then
collected in the same vial and stained free-floating under
identical conditions.
. All tissue
sections through the medial septal nucleus and vertical diagonal band
nucleus were saved, starting with 1.6 mm anterior to bregma
through 0.3 mm posterior to bregma (anatomical landmarks were
used to determine anterior-posterior level). One-third of these
tissue sections were immunostained for ChAT, one-third were
immunostained for VAChT, and one-third were immunostained for TrkA.
We also saved sections through the dorsal CA1 region of the
hippocampus from animals receiving infusions in that site. Tissue
sections from differently trained animals were nicked at a
predetermined place on the cortex (for later identification) and then
collected in the same vial and stained free-floating under
identical conditions.
Cholinergic neurons are known to coexpress ChAT and VAChT proteins (Schafer
et al., 1995![]() ; Gilmor et al., 1996
; Gilmor et al., 1996![]() ; Arvidsson et
al., 1997
; Arvidsson et
al., 1997![]() ; Ichikawa et al.,
1997
; Ichikawa et al.,
1997![]() ; Roghani et
al., 1998
; Roghani et
al., 1998![]() ; Schäfer
et al., 1998
; Schäfer
et al., 1998![]() ). We used antibodies
directed against TrkA, ChAT, and VAChT to stain the cholinergic
neurons of the medial septal area and the cholinergic terminals in
the hippocampus. To exclude effects caused by nonspecific axonal
damage, we also used antibodies to synaptophysin, which stain nerve
terminals nonselectively, on hippocampal sections.
). We used antibodies
directed against TrkA, ChAT, and VAChT to stain the cholinergic
neurons of the medial septal area and the cholinergic terminals in
the hippocampus. To exclude effects caused by nonspecific axonal
damage, we also used antibodies to synaptophysin, which stain nerve
terminals nonselectively, on hippocampal sections.
Anti-TrkA (polyclonal Ab 1577 purchased from Sigma, St. Louis, MO) was
diluted 1:500 in PBS containing 0.1% sodium azide. Anti-ChAT (goat
polyclonal Ab obtained from Chemicon, Temecula, CA) was diluted 1:200
in PBS with 0.1% sodium azide. Anti-VAChT (rabbit polyclonal Ab) (see
Roghani et al., 1998![]() ) and
anti-synaptophysin (monoclonal Ab, clone SVP-38, obtained from Sigma)
were both diluted 1:1000 in PBS with 0.1% sodium azide. After
24 hr of incubation at 4°C in the primary antibody, brain tissue
sections were transferred to biotinylated secondary antibodies and
then rinsed and exposed to a solution of avidin-biotin complex
(Vector Laboratories, Burlingame, CA). Finally, tissue sections were
incubated in a PBS solution containing 0.05% diaminobenzidine, 0.01%
hydrogen peroxide, and 2.5% nickel ammonium sulfate.
) and
anti-synaptophysin (monoclonal Ab, clone SVP-38, obtained from Sigma)
were both diluted 1:1000 in PBS with 0.1% sodium azide. After
24 hr of incubation at 4°C in the primary antibody, brain tissue
sections were transferred to biotinylated secondary antibodies and
then rinsed and exposed to a solution of avidin-biotin complex
(Vector Laboratories, Burlingame, CA). Finally, tissue sections were
incubated in a PBS solution containing 0.05% diaminobenzidine, 0.01%
hydrogen peroxide, and 2.5% nickel ammonium sulfate.
We analyzed these tissue sections with an Olympus microscope equipped with a
Bioquant image
analysis system. We assessed infusion sites in the following ways.
For animals infused in the medial septal area, we first examined the
tissue to establish the extent of protein disruption and then took
low-power photomicrographs of the affected regions. We verified that
the extent of protein disruption extended the same amount as did our
preliminary injections of fluorescein (FITC)-conjugated antisense
oligonucleotide provided by the manufacturer. For animals infused in
CA1 of the hippocampus, we mapped the extent of the infusion on
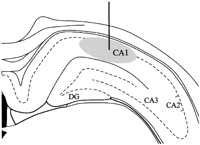
templates from the atlas of Paxinos and Watson (1997)![]() , as shown in Figure 1. We also
took high-power photomicrographs of cholinergic terminal loss at
the center of the infusion site.
, as shown in Figure 1. We also
took high-power photomicrographs of cholinergic terminal loss at
the center of the infusion site.
|
An unbiased sampling (video-assisted) technique was used to count and measure cross-sectional areas of synaptophysin terminals in CA1 of the hippocampus. On the basis of anatomical landmarks, the region of CA1 located 4.5 mm posterior and 3.0 mm lateral to bregma was selected with the microscope set at 10×. The lens was focused on the pyramidal cell layer (viewer assisted) and then moved into the immediately adjacent stratum radiatum (while the observer was not viewing the material under the microscope). The objective was then changed to 40×, and computer-assisted measurements were made. Slides from animals receiving different treatments were mixed together and assigned nondescriptive numbers before assessments were made such that the observer was blind to the training each animal received.
We also made analyses of remote effects in the medial septal area after
infusions of oligonucleotides into CA1 of the hippocampus. Because
ChAT and VAChT immunoreactivities completely fill cholinergic somata,
we were able to evaluate cholinergic cell cross-sectional area in
these immunostained sections. Cell body cross-sectional area
measurements were made for every cholinergic neuron throughout the
medial septal nucleus and vertical diagonal band nucleus in sections
immunostained for ChAT or VAChT after digitization of the images onto
a video screen. Cell fragments, identified by their irregular shape
and small cross-sectional area, were not included in the analyses.
The boundaries of these brain areas were taken from the atlas of
Paxinos and Watson (1997)![]() . The observer
was blind to the training each animal received. We attempted to
use fully the stereological method to attain measurements that
would enable us to estimate cell volume; however, we were cautious
in doing so because we had introduced 5-10 µl of fluid into each
hemisphere, thereby altering the volume of affected brain areas.
Nonetheless, we adhered to the principles of unbiased stereology
in every other way possible (e.g., in tissue sectioning and selection
of sections to analyze).
. The observer
was blind to the training each animal received. We attempted to
use fully the stereological method to attain measurements that
would enable us to estimate cell volume; however, we were cautious
in doing so because we had introduced 5-10 µl of fluid into each
hemisphere, thereby altering the volume of affected brain areas.
Nonetheless, we adhered to the principles of unbiased stereology
in every other way possible (e.g., in tissue sectioning and selection
of sections to analyze).
Statistics. We used a one-way ANOVA in most cases and a two-way ANOVA when specified. We also used the Student-Newman-Keuls post hoc test to assess differences in behavioral scores and NGF levels at different training-testing intervals and the Persson correlation coefficient to assess the significance of correlations between cell body cross-sectional area and behavioral responding.
| |
RESULTS |
|---|
Time course of behavioral and hippocampal NGF responses
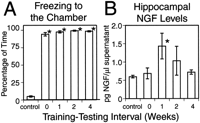
The rats acquired a high rate of the behavioral response freezing after two training sessions of tone paired with shock delivered in a novel chamber. As shown in Figure 2A, the percentage of time animals froze was consistently high and significantly above levels shown by naive controls (p < 0.01). Without training, naive animals showed little or no freezing when placed in the chamber (see Fig. 2A, control bar).
|
Although the behavioral response was maintained across the 4 week period we assessed, hippocampal NGF levels showed a distinct pattern of change after training (see Fig. 2B). Animals killed 24 hr after training (0 week) had NGF levels that were not significantly different from that of naive controls. A marked increase in NGF levels occurred, however, at 1 week after training. In these animals, NGF levels were 72% higher than those of naive controls, and this increase was significant (p < 0.05). NGF levels were not significantly elevated 2 weeks after training, and by 4 weeks after training, NGF levels in the hippocampus were indistinguishable from levels found in naive controls.
NGF quantitation in the parietotemporal cortex did not show the same pattern we found for hippocampal NGF. At 1 week after training, NGF in the parietotemporal cortex was found to decrease by 60% in trained animals as compared with naive controls (p < 0.05). This was the only time interval that revealed any differences between trained and control animals.
The effects of testing and the existence of any interactions between testing and training were also assessed by two-way ANOVA. To maximize effects of the test, rats were killed 1 hr after testing. At this short interval after testing, the effect of testing was significant (p < 0.01). Training produced nonsignificant trends in the same direction as described above (i.e., hippocampal NGF levels of trained rats were increased 31% above those of untrained rats, and NGF in the parietotemporal cortex was decreased by 47% in trained rats compared with untrained rats). There were no significant interactions between effects of training and testing shown by our two-way ANOVA.
Effects of antisense TrkA oligonucleotides on behavioral responses
We placed antisense oligonucleotides into CA1 of the hippocampus to cause
specific knockdown of cholinergic markers. This kind of specificity
was sought, in part, because lesions of that structure have produced
results that are open to varied interpretations (see Gewirtz et al.,
2000![]() ). These
infusions of antisense TrkA oligonucleotide produced changes in the
behavioral responses that depended on the unilateral versus bilateral
nature of the infusion, the site, and time interval after training
(see Fig. 3). Our
first group of animals received unilateral infusions of antisense
TrkA oligonucleotides into the medial septal area 1 week after
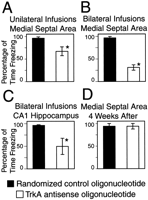
training. As shown in Figure 3A,
these animals froze significantly less when placed in the chamber
compared with animals given randomized oligonucleotides
(p < 0.05). As shown in Figure 3B,
bilateral infusions of antisense TrkA oligonucleotide decreased
freezing to an even greater extent than did unilateral infusions. The
level of significance for freezing differences between bilateral
infusion of antisense oligonucleotide and bilateral randomized
oligonucleotide was greater (p < 0.01) than this
comparison for unilateral infusions. This may reflect a dose-response
effect, because the bilateral infusions involved twice as much
oligonucleotide as did the unilateral infusions (a total of
20 µl compared with 10 µl).
). These
infusions of antisense TrkA oligonucleotide produced changes in the
behavioral responses that depended on the unilateral versus bilateral
nature of the infusion, the site, and time interval after training
(see Fig. 3). Our
first group of animals received unilateral infusions of antisense
TrkA oligonucleotides into the medial septal area 1 week after
training. As shown in Figure 3A,
these animals froze significantly less when placed in the chamber
compared with animals given randomized oligonucleotides
(p < 0.05). As shown in Figure 3B,
bilateral infusions of antisense TrkA oligonucleotide decreased
freezing to an even greater extent than did unilateral infusions. The
level of significance for freezing differences between bilateral
infusion of antisense oligonucleotide and bilateral randomized
oligonucleotide was greater (p < 0.01) than this
comparison for unilateral infusions. This may reflect a dose-response
effect, because the bilateral infusions involved twice as much
oligonucleotide as did the unilateral infusions (a total of
20 µl compared with 10 µl).
|
As shown in Figure 3C, antisense TrkA oligonucleotide also decreased freezing in animals receiving bilateral infusions into CA1 of the hippocampus compared with animals having randomized oligonucleotides infused into the same site (p < 0.05). When we used this infusion site, changes in freezing appeared to be caused by remote effects on the medial septal area because these alterations in behavior did not appear until 48 hr after the infusions were made. Animals with infusions into CA1 of the hippocampus did not show contextual memory impairments at 24 hr (data not shown), as did the animals with infusions into the medial septal area.
The sensitivity of the contextual memory to antisense TrkA oligonucleotides was time dependent. At 4 weeks after training, animals receiving infusions of antisense TrkA oligonucleotide into the medial septal area did not show decreased freezing compared with control animals given randomized oligonucleotide (Fig. 3D). Also, for all four groups of animals shown in Figure 3, freezing to tone was unaffected by the antisense TrkA oligonucleotide (data not shown). Hence the results of antisense TrkA oligonucleotide infusions into the hippocampus were specifically related to contextual memory consolidation.
Histochemical confirmation of antisense TrkA knockdown near the infusion site and remote effects on medial septal neurons after hippocampal infusions
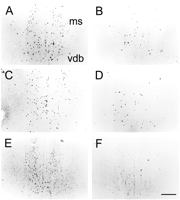
Antisense TrkA oligonucleotide substantially decreased cholinergic markers along the septohippocampal pathway. Unilateral and, to a greater extent, bilateral infusions of antisense TrkA oligonucleotides decreased immunostaining intensity in the medial septal area (i.e., the medial septal nucleus and vertical diagonal band nucleus). As shown in Figure 4, the number of cells showing detectable amounts of ChAT, VAChT, or TrkA in animals receiving randomized control oligonucleotide (A, C, E) was reduced by unilateral infusions of antisense TrkA oligonucleotide (B, D, F). Approximately 35% of cells in the medial septal area remained immunopositive for ChAT and VAChT, whereas only 20% remained immunopositive for TrkA, after unilateral infusions of antisense TrkA oligonucleotide as compared with randomized control oligonucleotide (Fig. 4, compare B,D,F; A,C,E). Hence the strongest effect of the antisense TrkA oligonucleotide was directly on TrkA receptor expression. Bilateral infusions produced reductions of >90% in the number of cells stained (data not shown).
|
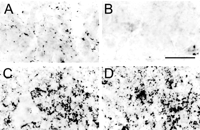
Infusions of antisense TrkA oligonucleotide into CA1 of the hippocampus caused local and remote effects. Near the infusion site, cholinergic terminal markers were reduced in density. As shown in Figure 5, the density and staining intensity of VAChT-containing terminals present in animals receiving randomized oligonucleotide (A) were visibly reduced after infusions of antisense TrkA oligonucleotide (B). The extent of reduced staining, as depicted in Figure 1, was limited to the CA1 region. The reduction in density and staining intensity was specific for the cholinergic markers VAChT (see above) and ChAT (data not shown) and was not observed in sections immunohistochemically processed with anti-synaptophysin, which stains all nerve terminals regardless of transmitter type (Fig. 5, compare C, D). Synaptophysin-stained terminals did not significantly differ between animals receiving TrkA oligonucleotide versus randomized oligonucleotide in terms of density (6.8 × 104 vs 5.6 × 104 terminals/mm2 for the left hippocampus and 6.4 × 104 vs 5.3 × 104 terminals/mm2 for the right hippocampus, respectively; p > 0.05) or area of terminal boutons (0.245 vs 0.258 µm2 for the left hippocampus and 0.290 vs 0.273 µm2 for the right hippocampus, respectively; p > 0.05). Our observation that synaptophysin-stained terminals were not significantly different between groups of rats receiving different treatments possibly excludes any other reason (e.g., stage of estrous cycle) from conceivably accounting for significant differences between behavioral and cellular measurements between groups.
|
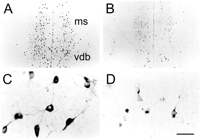
We also assessed the medial septal area for remote cellular changes after the infusions of antisense TrkA oligonucleotide into CA1 of the hippocampus. We further compared these remote changes, which were far removed from any damage at the infusion site, to the behavioral response rates. Although the entire hippocampus was not affected by our antisense TrkA infusions, cell bodies throughout the medial septal nucleus and the vertical diagonal band nucleus showed changes, suggesting that many of these cells retrogradely transported the antisense oligonucleotide from the CA1 region. As shown in Figure 6, the number of cells expressing ChAT in the medial septal nucleus and vertical diagonal band nucleus that were present after randomized oligonucleotide infusions (A) was visibly reduced after infusions of antisense TrkA oligonucleotide (B) into CA1 of the hippocampus. Moreover, the remaining ChAT-immunopositive cells had visibly reduced cell body cross-sectional areas (Fig. 6, compare D, C).
|
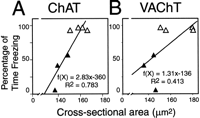
In our tissue samples, immunoreactive binding to the cytosolic proteins ChAT and VAChT was localized throughout the entire intracellular space; it filled each cell body completely, reaching the inner cell membrane and thereby enabling cell area measurements. Cell body cross-sectional area reductions were evident in analyses of both the ChAT- and the VAChT-immunostained sections. Pooling the data for animals receiving antisense TrkA oligonucleotide or randomized oligonucleotide, average cell body cross-sectional area for both ChAT- and VAChT-stained cells ranged from 129 to 181 µm2, and these average cross-sectional areas varied across animals primarily in accordance with the treatment received (see Fig. 7). In both series of tissue sections, the average cell body cross-sectional area for each animal correlated significantly with its behavioral response to the chamber. For ChAT cell body cross-sectional area and freezing, the correlation was more significant (p < 0.005) than that for VAChT cell body cross-sectional area and freezing (p < 0.05).
|
| |
DISCUSSION |
|---|
NGF alterations in the hippocampus at 1 week
The results from our first experiment show that NGF is temporarily increased during a critical period of contextual memory consolidation. An increase in hippocampal NGF levels, above that of naive controls, was only apparent 1 week after training. There was no increase above control levels the day after training, nor was there any sign of an increase at 2 or 4 weeks after training. Thus, changes in NGF levels reflect a short-lived, sensitive period for plastic change in the hippocampus, a time during which the permanent memory for a new spatial context is stored.
Because NGF is a regulator of neural plasticity, this increase in NGF level
might be expected to precede structural changes that accompany
contextual memory consolidation, such as those involving the
degradation of microtubule-associated protein-2 (MAP-2). Degradation
of MAP-2 occurs in the hippocampus 2 weeks after the
establishment of a conditioned fear response to the same spatial
context (Woolf et al., 1999![]() ). Thus, the
increased NGF level shown in the present study would come slightly
before the MAP-2 degradation shown previously.
). Thus, the
increased NGF level shown in the present study would come slightly
before the MAP-2 degradation shown previously.
Initially, we were concerned that changes in NGF levels could be affected by
mere exposure to the tone during testing. Simply removing rats from
their home cages and exposing them to tone increases c-fos expression
in many brain regions independent of conditioning (Smith et al.,
1992![]() ). Induction
of c-fos expression is maximal 1 hr after tone stimulation
(Zuschratter et al., 1995
). Induction
of c-fos expression is maximal 1 hr after tone stimulation
(Zuschratter et al., 1995![]() ). Because
c-fos protein is expressed in cells that respond to NGF (Sharp et
al., 1993
). Because
c-fos protein is expressed in cells that respond to NGF (Sharp et
al., 1993![]() ), increases
in NGF level might be anticipated up to 1 hr after testing. To
avoid this problem, we waited until 4 hr after testing to assess
for training-related differences in NGF level. No obvious effects of
testing were observed, and because all groups of animals were
similarly exposed to tone during testing, the specific NGF increase
at 1 week could not be attributed to mere exposure to the tone
or chamber during testing. In tissue assayed for NGF within 1 hr
after testing, the testing procedure, which consisted of exposure to
the training chamber and the tone, produced a large effect. We found
no interactions between training and testing in these animals,
however, indicating that the test did not intensify or elicit any
latent effects because of training in our first result. Hence we
confidently interpret our NGF increase at 1 week to be wholly
attributable to contextual memory consolidation occurring at this
time. Our results are also consistent with enhanced retention of
inhibitory avoidance behavior shown after NGF infusions into CA1 of
the hippocampus after training (Walz et al., 2000
), increases
in NGF level might be anticipated up to 1 hr after testing. To
avoid this problem, we waited until 4 hr after testing to assess
for training-related differences in NGF level. No obvious effects of
testing were observed, and because all groups of animals were
similarly exposed to tone during testing, the specific NGF increase
at 1 week could not be attributed to mere exposure to the tone
or chamber during testing. In tissue assayed for NGF within 1 hr
after testing, the testing procedure, which consisted of exposure to
the training chamber and the tone, produced a large effect. We found
no interactions between training and testing in these animals,
however, indicating that the test did not intensify or elicit any
latent effects because of training in our first result. Hence we
confidently interpret our NGF increase at 1 week to be wholly
attributable to contextual memory consolidation occurring at this
time. Our results are also consistent with enhanced retention of
inhibitory avoidance behavior shown after NGF infusions into CA1 of
the hippocampus after training (Walz et al., 2000![]() ).
).
TrkA knockdown decreases freezing responses elicited by the context
Low levels of NGF precluded our NFG assay being limited to any subregion of the hippocampus; however, our antisense TrkA infusions were specific to CA1 of dorsal hippocampus. These TrkA knockdown experiments confirmed that increased hippocampal NGF present at 1 week after training is specifically involved with consolidation of contextual memory to the extent that freezing to context was impaired but freezing to tone was unimpaired. At 4 weeks after training, however, the contextual memory appeared to be fully consolidated, and antisense TrkA infusions into the medial septal area did not produce any deficit in freezing.
Although waiting 1 week after training produced the same behavioral deficit regardless of infusion site, the time after which surgical infusion caused behavioral deficits was site dependent. For the medial septal area, behavioral response was diminished 24 hr after infusion. Antisense TrkA oligonucleotide infused into CA1 hippocampus, however, produced effects on behavioral response after 48 hr but not after 24 hr. This could be because extra time was needed for the antisense TrkA oligonucleotide to be retrogradely transported to the cell bodies of the cholinergic neurons, which are located in the medial septal area. After retrograde transport to medial septal cell bodies, one would expect reduced expression of cholinergic proteins and a behavioral deficit.
It is also worth mentioning that a very low level of TrkA mRNA has been
detected in CA1 of the hippocampus by use of in situ
hybridization with a sensitive P33-labeled riboprobe, and its
presence has been confirmed using reverse transcription-PCR
(Cellerino, 1996![]() ). Antisense TrkA
oligonucleotide decreased the staining intensity of terminals in CA1
of the hippocampus in our experiment. This might represent a
withdrawal of cholinergic terminals, which has been described
previously for a TrkA antagonist (Debeir et al., 1999
). Antisense TrkA
oligonucleotide decreased the staining intensity of terminals in CA1
of the hippocampus in our experiment. This might represent a
withdrawal of cholinergic terminals, which has been described
previously for a TrkA antagonist (Debeir et al., 1999![]() ). A withdrawal of
cholinergic terminals in the hippocampus might be expected to produce
behavioral consequences. It is also possible that behavioral effects
are mainly attributable to retrograde effects on medial septal
neurons. This latter premise is supported by the time delay and the
significant correlations between ChAT cell body cross-sectional area
and behavioral response rate. It is also possible that local effects
in CA1 of the hippocampus and remote effects in the medial septal
area both contribute to diminished behavioral responses. Hence the
antisense knockdown of the TrkA receptor could have inhibited two
functionally important sites simultaneously.
). A withdrawal of
cholinergic terminals in the hippocampus might be expected to produce
behavioral consequences. It is also possible that behavioral effects
are mainly attributable to retrograde effects on medial septal
neurons. This latter premise is supported by the time delay and the
significant correlations between ChAT cell body cross-sectional area
and behavioral response rate. It is also possible that local effects
in CA1 of the hippocampus and remote effects in the medial septal
area both contribute to diminished behavioral responses. Hence the
antisense knockdown of the TrkA receptor could have inhibited two
functionally important sites simultaneously.
TrkA knockdown affects cholinergic markers ChAT and VAChT
Our data suggest that NGF acted via the TrkA receptor to regulate cholinergic markers ChAT and VAChT during the sensitive period for contextual memory consolidation occurring 1 week after training. We directly confirmed via immunohistochemical detection that the antisense TrkA oligonucleotide markedly decreased TrkA protein, as well as ChAT and VAChT proteins, in cholinergic cells of the medial septal area after infusions into that site. Previous studies have suggested that NGF may participate in memory consolidation via the upregulation of ChAT (see introductory remarks). Accordingly, the increased hippocampal NGF we found during contextual memory formation would be expected to upregulate ChAT gene expression, as well as that of the closely linked VAChT gene. Moreover, synaptophysin immunoreactivity in hippocampal sections was unaltered, confirming that TrkA knockdown of ChAT and VAChT proteins, and not general axonal damage, was responsible for the memory deficit.
Morphological and histochemical correlates of contextual memory
A number of studies have reported changes in cholinergic cell body
cross-sectional area that correlate with learning, memory, or
exposure to an enriched environment. One of the initial reports was
from our laboratory in which we reported increased cell body
cross-sectional area after the acquisition of a conditioned emotion
response to tone demonstrated by in situ hybridization for
ChAT mRNA (Oh et al., 1996![]() ). Gustilo et al.
(1999)
). Gustilo et al.
(1999)![]() reported
recently that NGF increases cholinergic cell body area, and this
increase may account for NGF-induced enhancement of recent and, to
some extent, reference memory. Nonetheless, these authors
importantly caution that corresponding effects on cholinergic
terminals may be more critical to memory function than cell body
alterations. Cell body area increases, however, have also been
reported for TrkA- and p75-immunostained cholinergic cells in the
medial septal area after a year-long exposure to an enriched
environment (Pham et al., 1999a
reported
recently that NGF increases cholinergic cell body area, and this
increase may account for NGF-induced enhancement of recent and, to
some extent, reference memory. Nonetheless, these authors
importantly caution that corresponding effects on cholinergic
terminals may be more critical to memory function than cell body
alterations. Cell body area increases, however, have also been
reported for TrkA- and p75-immunostained cholinergic cells in the
medial septal area after a year-long exposure to an enriched
environment (Pham et al., 1999a![]() ).
).
All the above studies reported increased cell body area under enhanced conditions. In this report, we present evidence that antisense to TrkA infused into CA1 of the hippocampus decreases the cell body cross-sectional area of medial septal neurons, and moreover, these cell cross-sectional areas are correlated with behavioral measures on the contextual memory task. The present data are highly compatible with previous results and warrant serious consideration of the basic tenant that not only do cholinergic cell bodies show increases in cross-sectional area during memory consolidation but these cross-sectional area increases are somehow necessary to the memory consolidation processes.
Although alterations in cell body area have now been repeatedly documented,
effects on cholinergic terminals may be equally (or more) relevant to
memory function. Cell body cross-sectional area alterations might
signify, for example, reorganized trafficking to cholinergic
terminals that will affect neurotransmitter release for extended
periods of time. We were able to document dramatic local effects on
VAChT terminals after antisense TrkA oligonucleotide was infused into
CA1 of the hippocampus. It seems probable that cholinergic terminals
are structurally altered shortly before the recipient cholinoceptive
cells in the hippocampus undergo structural modification (see Woolf,
1996a![]() ,b
,b![]() , 1998
, 1998![]() ). We plan to
undertake more detailed studies relating the effects that antisense
oligonucleotides have on cholinergic terminals in the hippocampus
and correlative changes in behavioral response.
). We plan to
undertake more detailed studies relating the effects that antisense
oligonucleotides have on cholinergic terminals in the hippocampus
and correlative changes in behavioral response.
The present results demonstrating involvement of ChAT and VAChT proteins in
the memory consolidation process may be interpreted as implying that
changes in ACh synthesis and release are a part of this NGF-mediated
process. In a previous study, NGF treatment of primary cultured rat
embryonic septal cells enhanced expression of VAChT and ChAT
proteins, as well as increased ACh synthesis and release (Oosawa et
al., 1999![]() ). Thus, it is
reasonable to assume that the memory impairment we observed in
animals infused with the antisense TrkA oligonucleotide was caused,
in part, by a reduction in synthesis and release of ACh by the
presynaptic terminals in the CA1 of hippocampus.
). Thus, it is
reasonable to assume that the memory impairment we observed in
animals infused with the antisense TrkA oligonucleotide was caused,
in part, by a reduction in synthesis and release of ACh by the
presynaptic terminals in the CA1 of hippocampus.
In summary, these results support significant involvement of receptor-mediated mechanisms in conditioned behavior. Studying the consequences of such neuronal mechanisms should contribute to our understanding of the role of synaptic transmission by neurotransmitters, such as ACh, in normal complex brain functions as well as in age-related neurological disorders.
| |
FOOTNOTES |
|---|
Received Sept. 15, 2000; revised Nov. 1, 2000; accepted Nov. 14, 2000.
This work was supported by National Institutes of Health Grant NS 35656 to A.R. We thank Shazia Iqbal for her contributions.
Correspondence should be addressed to Dr. Nancy J. Woolf, Department of Psychology, University of California, Los Angeles, CA 90095-1563. E-mail: nwoolf@ucla.edu.
| |
REFERENCES |
|---|
|
|